| Einführung | Raubtieranatomie | Stammbaum | Molekunargenetik |
| Mutation & Varietät | Genetische Verarmung | Referenzen |
4. Molekulargenetik
4.1. Die molekulare Ära der Biologie
In den 1950er Jahren begann die Aufklärung der dreidimensionalen Struktur von
Proteinen und, etwa zwanzig Jahre später, der routinemässigen Ermittlung von
DNA-Sequenzen (die englische Abkürzung DNA statt DNS ist gebräuchlich). Obwohl
es vielleicht einseitig erscheint, kann man hier vom Beginn einer "molekulare
Ära" der Biologie sprechen. Die Erfassung der Gensequenz-Daten hat sich seither
ständig beschleunigt, und die entsprechenden Datenbanken sind mit wenigen
Ausnahmen frei zugänglich. Damit wurden erstmals auch Vergleiche der DNA-Sequenzen
verschiedener Tierstämme bis hinunter zu verschiedenen Individuen derselben Art
möglich und man begann, den so messbaren Abstand in der evolutionären Entwicklung
zu quantifizieren und für Verwandtschaftsbestimmungen zu nutzen. Inzwischen
werden Methoden der Molekulargenetik immer häufiger eingesetzt.
Die Terminologie der Molekularbiologie und Reproduktionsmedizin führt allerdings
oft zu einiger Verwirrung. Nicht selten ist es erst die grobe Vereinfachung der
Sachverhalte in der Presse, die zu Missverständnissen führt. Oft werden auch zu
grosse Erwartungen geweckt, die sich später nicht einlösen lassen, weil z.B.
eine neuentdeckte Genfunktion nicht monokausal für eine bestimmte Erbkrankheit
verantwortlich ist. Auch wird beim ermitteln von DNA-Sequenzen im Zusammenhang
mit dem "Human Genome Projekt" manchmal von "Bauplänen" gesprochen, die man mit
gentechnischen Methoden erhalte. Dies würde implizieren, dass man Bakterien oder
gar höhere Organismen anhand der DNA-Sequenz im Labor nachbauen könnte.
Dies ist natürlich nicht möglich, weil die Kenntnis über die Zusammensetzung von
Einzelteilen (Proteine) noch keine Konstruktionsanleitung für den Gesamtorganismus
darstellt. Vergleichbar ist die Behauptung, ein Mensch würde aus chemischen
Substanzen bestehen, die in der Apotheke für den Preis eines Mittagessens zu
erwerben wären. Das ist zwar kaufmännisch korrekt, aber auch völlig irrelevant,
weil es keine Maschine gibt, die einen Menschen aus den preiswerten Grundsubstanzen
zusammenbauen könnte. Ein ebenfalls oft gesehenes Beispiel ist die Vermischung von
Themen der Reproduktionsmedizin und Gentechnik im Begriff "Klonen". Der Begriff der
"Chimäre" hat ebenfalls verschiedene Bedeutungen, wobei der Ausdruck in der Biologie
genau definiert ist (siehe Kap. 5).
4.2. Proteinevolution
Proteine bestehen aus verschiedenen Ketten von Aminosäuren und die Abfolge dieser
Aminosäuren, und damit schliesslich die 3-dimensionale Form der Proteine, wird
durch die DNA-Sequenz im Zellkern festgelegt. Die DNA selbst verbleibt im Zellkern
während der Lebensspanne der Zelle. Wenn ein bestimmtes Protein hergestellt werden
muss, werden Mechanismen aktiviert, die eine RNA-Kopie von spezifischen Sequenzen
der DNA herstellen (messenger-RNA) und aus dem Zellkern herausbringen.
Danach wird diese messenger-RNA von Ribosomen abgelesen und ein Protein wird nach
der vorliegenden Sequenzinformation hergestellt. Der Plan zu diesem Protein ist im
Zellkern verpackt und wird dort bei der Zellteilung dupliziert, mit der Ausnahme
der mitochondrialen DNA (siehe unten). Die Sequenz der DNA oder der RNA wird als
Basensequenz geschrieben, d.h. die Codierung der Erbinformation in der DNA-Doppelhelix
besteht aus der Abfolge von 4 Pyrimidin- bzw. Purin-Basen (Adenin, Thymin,
Guanin, Cytosin) und kann deshalb als eine Buchstabenkette wie z.B. "ACGAAT...."
geschrieben werden. Die Erbinformation eines Menschen ist mehrere Milliarden
Buchstaben bzw. Basen lang.
Ein Gen ist ein verschieden langer Abschnitt der DNA, der auch unterbrochen sein
kann und die Codierung für eine Polypeptidkette (=Protein) oder eine RNA-Sequenz
sowie Steuersequenzen enthält. Bei einfacheren Lebewesen (Bakterien) codiert 1
Gen genau für 1 Protein. Bei höheren Lebewesen entstehen aus einem Gen jedoch
mehrere Proteine, weshalb die Gesamtzahl der Gene beim Menschen auch nicht mehr
als 30'000-40'000 beträgt, wie wir seit dem Abschluss des "Human Genome Projects"
nun sicher wissen.
Gene und die daraus entstehenden Proteine verändern sich im Laufe der Geschichte.
Da wir die Chemische Natur der Erbsubstanz kennen, können wir den Begriff der
Mutation genauer, als eine Veränderung der Sequenz/Abfolge oder Anzahl der Basen
der DNA, definieren.
Es zeigte sich auch, dass sich verschiedene Proteine mit unterschiedlicher
Geschwindigkeit verändern. Eine naheliegende Erklärung liegt darin, dass Proteine
Bereiche enthalten, die für seine jeweilige Funktion mehr oder weniger lebenswichtig
sind. Ein Enzym, das z.B. an der Đbertragung von Nervensignalen bei Synapsen beteiligt
ist, besitzt ein verarbeitendes Zentrum, an dem die eigentliche Verarbeitung der
chemischen Substrate stattfinden und darüber hinaus periphere Strukturen. Eine
Mutation am sog. aktiven Zentrum hat eine sofortige Änderung in den Eigenschaften
des Proteins zur Folge (langsamere Verarbeitung, schneller, gar nicht, andere
Vorlieben für bestimmte Substrate etc.). Fällt diese Mutation schlussendlich im
Verbund des Körpers zum Nachteil der Fortpflanzung desjenigen Lebewesens aus, das
die Mutation trägt, so wird die veränderte Erbinformation nicht weitervererbt.
Daher verändert sich das Protein im Laufe der Evolution an der Stelle, die für
das Zentrum codiert, langsamer.
Auf der tieferen Ebene der DNA-Sequenz kommt noch dazu, dass es viele verstreute
Bereiche gibt, die keine sinnvolle Informationen enthalten. Weil für diese
Bereiche auch kein Selektionsdruck herrscht, verändern sie sich öfter. Da nun
also die Evolutionsrate der Proteine verschieden ist, kann man beim Vergleich von
zwei oder mehreren Tierarten ein Protein wählen, dass sich in einer Geschwindigkeit
verändert, die für eine gegebene Fragestellung passend ist. Ist die Frage nach der
Verwandtschaft innerhalb nahe verwandten Arten, wie z.B. zwischen Mensch und
Schimpanse oder zwischen verschiedenen Gruppen von Menschen, so wird man Gene
auswählen, die sich rasch verändern.
4.3. DNA-Fingerabdruck und Verwandtschaftsbestimmung
Ein DNA-Fingerabdruck ist ein Nachweis von DNA-Sequenzen, die im hier gezeigten
Beispiel eines Vaterschaftstests nach den Mendelschen Regeln vererbt werden. Es
wird dabei nicht die ganze DNA eines Lebewesens miteinander verglichen, sondern
nur gezielt ausgewählte Sequenzen, also kurze Stücke des genetischen Textes
(siehe unten). Diese kurzen Stücke kommen in der ganzen Erbsubstanz viele
tausende Male vor und wandern zusammen in einem elektrischen Feld. In der Praxis
erhält man im Labor ein Bild eines Bandenmusters. Diese Bilder werden meist auf
einem Röntgenfilm entwickelt und lassen sich leicht mit einem anderen Bandenmuster
vergleichen. Ähnliche Methoden werden für die Verwandtschaftsbestimmung von
Wildtieren verwendet.
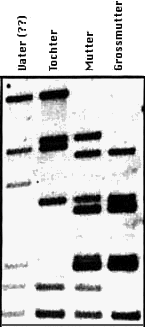
Ein einfaches Beispiel eines Vaterschaftstests (siehe Abbildung links): DNA vom
vermuteten Vater, von der Mutter, Tochter und Grossmutter mütterlicherseits wurde
gewonnen, zerkleinert, elektrophoretisch aufgetrennt und auf die aus drei Basen
bestehende Sequenz "CAC" getestet, wobei Fragmente verschiedener Länge = dunkle
Banden verschiedener Position in der Vertikalen entstanden sind.
Jedes Elternteil gibt etwa die Hälfte der Fingerprint-Banden an das Kind weiter,
wobei die vom jeweiligen Elternteil stammenden Banden eine zufällige Auswahl
aller Fingerprint Banden darstellen. Die Vaterschaft wurde in diesem Beispiel
zweifelsfrei bestätigt. Ein Nicht-Vater würde kaum eine einzige mit dem Kind
übereinstimmende Bande erzielen. Alle Banden des Kindes erscheinen entweder beim
Vater oder bei der Mutter. Dieser Test zeigt nur die Verwandtschaftsverhältnisse
auf. Er sagt nichts über die tatsächlich hervortretenden Eigenschaften aus.
(Bildquelle 3)
4.4. hypervariable mikrosatelliten-DNA
Sogenannte Mikro- oder Minisatelliten-DNA bezeichnet Sequenzwiederholungen in der
DNA. Dies sind kurze Abschnitte wie z.B. "TAACCC", die sich hintereinander (daher
auch 'Tandem-Wiederholungen') aufreihen und für die oft keine Funktion in der
Codierung für eine Protein bekannt ist. Bei den eigentlichen Minisatelliten-Sequenzen
findet man einen Polymorphismus (unterschiedliches Muster) von Individuum zu Individuum,
weshalb man anhand von Proben (z.B. ein Blutstropfen) Träger eines bestimmten Musters
identifizieren kann.
Der Unterschied von Probe zu Probe besteht nicht im Wortlaut der wiederholten
Sequenz, sondern in der Anzahl Wiederholungen, was offensichtlich keine biologischen
Konsequenzen zur Folge hat. In der Populationsgenetik kann man nun Proben von
verschiedenen Tierarten oder Gruppen der selben Art auf eine oder mehrere Minisatelliten
testen und erhält DNA-Fingerabdrücke. Diese lassen sich wiederum so auswerten, dass
eine Aussage über die Variabilität und die genetische Vielfalt der verschiedenen
Tierpopulationen gemacht werden kann (Harley et al, 2000). Da die Tandemwiederholungen
nur einem schwachen Selektionsdruck ausgesetzt sind, verändern sie sich im Laufe der
Abfolge von Generationen von Lebewesen rasch und sind deshalb "hypervariabel".
4.5. mitochondriale DNA (mtDNA)
Mitochondrien sind die kleinen Kraftwerke der Zellen. Sie sind zelluläre Körperchen
(Organellen), die ein wenig eigene DNA besitzen, was darauf hindeutet, dass sie
ursprünglich eine "freies" Leben ausserhalb der Körperzellen als bakterienähnliche
Lebensformen führten und bereits in einer frühen Phase der Evolution in die Zellen
der eukaryontischen Organismen aufgenommen wurden. Die DNA der Mitochondrien (mtDNA)
beinhaltet nur sehr wenige Gene, im Vergleich zur DNA im Zellkern. Konkret handelt
es sich dabei um 13 Proteine, die eine Rolle in der Energieproduktion der Mitochondrien
(Zellatmung) spielen, sowie 22 Transfer-RNA's und 2 ribosomale Proteine. Die
Mitochondrien selbst enthalten in ihrer Struktur viele zusätzliche Proteine, die aber
in der DNA des Zellkerns und nicht in den Mitochondrien selbst codiert sind.
Es gibt zwei wichtige Eigenschaften der mitochondrialen Erbsubstanz und ihrer
Vererbung:
-
Die mtDNA wird immer von der Mutter weitervererbt, weil bei der Verschmelzung von Eizelle und Spermium nur die Mitochondrien der Eizelle, also diejenigen der Mutter, übernommen werden. Weil die Vererbung der rein mitochondrialen Gene nicht über Chromosomen abläuft, gelten auch andere Regeln der Vererbung als die Mendel'schen. Veränderungen in der mtDNA stammen demzufolge nicht aus der Vermischung von Genen bei der Vererbung, sondern von Mutationen, also von Änderungen, die sich meist beim Kopieren der DNA einschleichen. Die Rate, mit der sich die Sequenzen zweier mtDNA's voneinander unterscheiden, liefert deshalb ein Mass für die Zeit, die vergangen ist, seitdem sich die beiden Stammbäume voneinander trennten.
-
Die mtDNA verändert sich rascher als diejenige im Zellkern (etwa 10 mal schneller). Eine Theorie dazu besagt, dass die biochemischen Bedingungen in den Mitochondrien bei der Energieproduktion die Entstehung von Mutationen begünstigt (Michikawa et al., 1999).
Die Veränderung der mtDNA im Laufe der Zeit kann als Mass für den Verwandtschaftsgrad
verschiedener Populationen von Lebewesen herangezogen werden. Da die Mutationsrate
der mtDNA hoch ist, können auch Voraussagen über kürzere Zeiträume (Jahrtausende)
getroffen werden. Wie bei anderen Methoden der Altersbestimmung muss mit
statistischen Modellen gearbeitet werden um eine Eichung der Werte zu erhalten.
Alle Gattungen der Familie der Felidae wurden bereits 1995 auf ihre
Verwandtschaftsverhältnisse mittels der Sequenzierung der mtDNA untersucht
(Janczewski et al, 1995). Dabei wurde der Stammbaum, so wie im Kapitel 3.1.2 gezeigt,
bestätigt. Diese Resultate gelten rückblickend für die Entwicklung der letzten 10
Millionen Jahre, da bisher nur mtDNA von heute lebenden Arten verwendet werden konnte.
Für andere Fragestellungen der Paläontologie wurde mtDNA aus fossilen Ueberresten
extrahiert und für Verwandtschaftsanalysen verwendet.
Je nach Fragestellung kann der Vergleich von DNA aus Mitochondrien oder aus dem
viel grösseren Anteil der DNA aus dem Zellkern zu verschiedenen Ergebnissen führen.
Mit der Weiterentwicklung dieses noch jungen Zweiges der Forschung wird man hier
in Zukunft noch mehr Erfahrungen sammeln müssen.
4.6. Variabilität des Haupt-Histokompatibilitäts-Komplex (MHC)
Die sog. MHC-Moleküle (für "major histocompatibility complex", MHC) waren schon
lange bekannt, bevor man ihre eigentliche Funktion im Immunsystem (auf die ich
hier nicht weiter eingehe) richtig einschätzen konnte. Die MHC-Moleküle sind die
zentralen Zielstrukturen bei Transplantations-Reaktionen. Wenn man Gewebe auf ein
Individuum derselben Art (Allo-Transplantation) oder auf ein Individuum einer
anderen Art (Xeno-Transplantation) überträgt wird es gewöhnlich abgestossen.
Diese Abstossung beruht auf einer Immunreaktion gegen eine bestimmte Gruppe von
Proteinen auf der Oberfläche der transplantierten Organe bzw. Hautstücke. Die
meisten dieser Proteine gehören zur MHC-Gruppe, weshalb der MHC hauptsächlich die
Abstossung oder Verträglichkeit von transplantierten Zellen und Gewebe bestimmt.
Die Eigenschaft, die für die Evolutionsbiologie von grossen Interesse ist, besteht
im sehr starken Polymorphismus der Gene, die für die MHC-Moleküle codieren. Mit
Polymorphismus ist gemeint, dass z.B. beim Menschen bis zu hundert verschiedene
Varianten der MHC-Gene bekannt sind, die wiederum an 5 verschiedenen Stellen im
Genom zu finden sind. Jedes menschliche oder tierische Individuum, ausser eineigen
Zwillingen hat somit ein eindeutiges MHC-Muster. Auf der Ebene einer Population oder
Gruppe ist die Variabilität um so grösser, je mehr Durchmischung mit anderen
Populationen stattfindet. Die MHC-Variabilität oder -Polymorphismus wurde deshalb
als Mass für die genetische Durchmischung einer bestimmten Gruppe von Wildtieren
aufgefasst und in einer Versuchsreihe an aus der Natur entnommenen Geparden mittels
Hauttransplantationen getestet [O'Brien et al, 1983] (siehe Kapitel 8)
4.7. Missverständnisse um einen alten Begriff: Chimären
Das Wort Chimäre bezeichnet ein Wesen, dessen Körper aus Teilen anderer Lebewesen
besteht. Die Chimaira der griechischen Mythologie war nach Homer ein feuerspeiendes
Ungeheuer, das Landschaften in Lykien verwüstete, bis es von Bellepheron mit Hilfe des
Flügelrosses Pegasus zur Strecke gebracht wurde. Die Chimäre war vorne Löwe, in der
Mitte Ziege und hinten Drache. Andere Mischwesen in der griechischen Mythologie sind
die Zentauren, Pferdekörper mit menschlichem Oberkörper und der Minotaurus, ein
menschlicher Körper mit Stierkopf. In der Mythologie der alten Griechen dienten die
Chimären, um Wesen zu beschreiben, die menschliche Eigenschaften aber gleichzeitig
auch die übermenschlichen Fähigkeiten von Tieren (Kraft, Schnelligkeit, Ausdauer)
besassen. Bis in die heutige Zeit finden Mischwesen (heutzutage meistens aus Mensch
und Maschine bestehend: "Cyborgs") Verwendung in phantastischen Erzählungen und in
der Populärkultur.
In der Biologie bezeichnet der Begriff Chimäre definitionsgemäss Individuen, die
aus genetisch unterschiedlichen Geweben bestehen oder genauer (nach Hentschel und
Wagner, Zoologisches Wörterbuch, 3. Auflage): "Komplexindividuum, das aus idiotypisch
verschiedenen Zellen besteht". Der Typ von Chimäre, der im Zusammenhang dieses Artikels
gemeint ist, entspricht der Vorstellung eines Mischwesens das sichtbar verschiedene
Tierarten in sich vereinigt, sodass die einzelnen Körperteile (Fell, Hörner, Kopf
und Körper etc.) weiterhin ihre ursprüngliche Form beibehalten.
Eine Gen-Chimäre ist ein genverändertes Tier, das in seinem Körper zwei oder mehrere
Gensequenzen verschiedener Herkunft in sich vereinigt, was sich in der äusseren
Gestalt des Tieres nicht unbedingt zeigen muss. Bei diesem zweiten Typ sind die
Begriffe transgenes oder genverändertes Tier gebräuchlicher. Transgene Tiere sind
in einem makroskopischen Sinn keine Mischwesen, da lediglich die Erbinformation
für einzelne Proteine des Körpers ausgetauscht oder verändert wurden.
Was den einen oder anderen Leser überraschen mag, ist die Tatsache, dass die am
besten bekannten und auch in Labors und Zuchtanstalten existierenden Chimären,
wie etwa die Schiege (aus Schaf und Ziege, engl. "geep") nicht durch Genmanipulation
entstehen, sondern lediglich durch das Mischen von ganz frühen Embryonen beider Arten
und darauffolgendem Wiedereinsetzen des gemischten Embryos, der erst aus wenigen
Zellen besteht, in ein Leihmutter-Tier. Diese Technik ist Teil der Fortpflanzungsmedizin,
was handwerklich anspruchsvoll ist und einige biochemische Kniffe erfordert, jedoch
das Erbmaterial völlig unberührt und unvermischt belässt. Die Mischung der Eigenschaften
dieser Chimären werden also nicht an etwaige Nachkommen weitergegeben!
Zumal die meisten dieser Chimären ohnehin steril sind, weil oft nicht nur z.B.
das Fell, sondern auch die Geschlechtsorgane ein Mosaik darstellen und nicht voll
funktionsfähig sind. Falls es doch Nachwuchs aus einer Paarung von Schiege/Schiege
oder einer Schiege mit einem Schaf oder einer Ziege gibt, ist das Junge doch wieder
nur entweder reines Schaf oder Ziege, weil die Geschlechtszellen der Schiege nicht
genetisch verschmolzen sind, sondern als heterogene Teile nebeneinander existieren.
Die grössten technischen Probleme, abgesehen von den erheblichen ethischen Bedenken,
bei der "Herstellung" einer Chimäre wie z.B. der Schiege liegen aber im Ablauf der
Schwangerschaft und im Anwachsen des Embryos in der Gebärmutter des Leih-Muttertiers.
Der Grund liegt in der Hülle bzw. Haut des gemischten Embryos, die oft nicht mit der
Gebärmutterwand der Leihmutter verträglich ist. Diese Abstossung ist ähnlich einer
Immunreaktion; es spielen dabei Oberflächenproteine der Zellen eine Rolle, die sich
von Art zu Art unterscheiden.
Dies ist auch der Grund, weshalb für die Chimären immer nur nahe verwandte Tierarten
oder genetisch veränderte Tiere derselben Tierart verwendet werden, wie Schaf und
Ziegen, bei denen es sehr selten sogar natürlicherweise zu "Bastarden" kommt. Die
Erbinformation der Hybriden ist dann tatsächlich vermischt, was noch nicht heisst,
dass die Tiere auch fruchtbar wären. Technisch gut etabliert ist die Erzeugung von
Chimären verschiedener Inzuchtstämme von Labormäusen mit jeweils unterschiedlicher
Fellfarbe, die dann z.B. an einem schwarz-weiss gefleckten Fell erkenntlich sind.
Dabei geht es aber um genetische Experimente innerhalb derselben Art (Maus), und
nicht um das Vermischen von Tierarten.
Eine weitere Hürde für die Entstehung von Hybriden von verschiedenen Arten bildet
die Anzahl Chromosomen, die z.B. bereits bei Mensch und Schimpanse, aber auch bei
Hund und Katze unterschiedlich ist. Die Anzahl Chromosomen ist nicht proportional
zur Entwicklungshöhe einer Art, sondern repräsentiert lediglich die "DNA-Verpackung"
der jeweiligen Arten. Siehe dazu die untenstehende Tabelle der Anzahl Chromosomen im
doppelten Chromosomensatz http://www.biokurs.de/skripten/13/bs13-1.htm.
Als Erklärung für die unterschiedliche Anzahl Chromosomen von Mensch und Schimpanse
wird eine Fusion zweier Chromosomen zu einem einzigen bei einem Vorfahren beider
Gattungen angenommen (IJdo et al. 1991, Yunis et al., 1980)
| lat. Name | dt. Name | Anzahl Chromosomen | |||
| Homo sapiens | Mensch | 46 | |||
| Pan troglodytes | Schimpanse | 48 | |||
| Canis sp. | Haushund | 78 | |||
| Felis cattus | Hauskatze | 38 | |||
| Solanum tuberosum | Kartoffel | 48 |
Die zellulären Mechanismen der Zellteilung würden bei unterschiedlicher Chromosomenzahl
bei der Befruchtung einer Eizelle durch ein Spermium einer anderen Tierart gestört und
die Folge wäre wiederum die Abstossung bzw. das Absterben des nicht lebensfähigen
Embryos in einem sehr frühen Stadium. Wie fast immer in der Biologie gibt es Spezialfälle,
wo die Toleranz des Systems scheinbar grösser ist: Manchmal kommt es bei der Zellteilung
der befruchteten Eizelle zu Chromosomen-Trennungsfehler in der sog. Reifeteilung. Das
Ergebnis sind Chromosomen-Aberrationen die sich durch eine ungewöhnlichen Anzahl
Chromosomenzahl im betroffenen Individuum auszeichnen. Manchmal sind diese Individuen
dennoch fruchtbar, aber beachten sie bitte, es handelt sich hier um Fortpflanzung
innerhalb der selben Art oder Varietät.
Wäre es nicht möglich, in einer Eizelle die Hälfte der DNA (oder noch einfacher:
die Hälfte der Chromosomen) des einen Tieres und die andere Hälfte eines anderen
Tieres zusammenzufügen? Dieses Experiment hätte wiederum kein lebensfähiges Tier
zur Folge. Verwandte oder in mehreren Kopien vorhandene Gene können gehäuft
nebeneinander auf demselben Chromosom liegen oder auf verschiedenen Chromosomen
verteilt sein. Die verschiedenen Gene liegen bei verschiedenen Tierarten nicht auf
denselben Chromosomen, sodass eine einfache Zusammenstellung eines gemischten
Chromosomensatzes in einem Verlust bzw. Vervielfältigung von Genen resultieren würde.
Ebenfalls darf man nicht vergessen, dass sich die Embryonen von verschiedenen
Tierarten unterschiedlich schnell und zu ebenso unterschiedlicher Grösse entwickeln.
Falls die Tierarten nicht sehr nahe verwandt sind, führt dies unweigerlich zu
Entwicklungsstörungen und zu einem sehr frühen Abbruch der Entwicklung des Embryos.
Im Extremfall sind in der Mythologie Chimären 'beschrieben' worden, von denen der
eine Anteil eine Entwicklung in einem Ei beenden würde, und der andere Anteil in
einer Gebärmutter (Vögel-Menschen/Löwen etc.). An dieser Stelle wäre die vollständige
Kontrolle über die Embryonalentwicklung eines Lebewesens nötig. An einen künstlichen
Uterus für Säugetiere in Form einer Maschine ist beim derzeitigen Erkenntnisstand
überhaupt nicht zu denken (es ist ja bisher noch nicht einmal möglich, künstliches
Blut herzustellen, das alle Eigenschaften des natürlichen Blutes aufweist).
Kaum einer Erwähnung bedarf die Tatsache, dass sich verschiedene Tierfamilien in
physiologischen Parametern wie Körpertemperatur, Herzfrequenz, Atmungsvolumen,
Verdauung und Nahrungs-Spezialisierung, Knochenstruktur sowie Versorgung und Steuerung
des Körpers mit Nervenimpulsen so stark unterscheiden, dass eine Chimäre im Stile
eines Minotaurus oder altägyptischer Götter mit Tierhäuptern nicht lange lebensfähig
wäre. In der Tat sind die physiologischen und biochemischen Unterschiede bei
verschiedenen Tieren so ausgelegt, dass z.B. das massive Vergrössern oder Verkleinern
eines Organs bzw. von deren Knochenstrukturen (Flügel, Kopf etc.) für eine phantastische
Chimäre eine völlige Neukonstruktion seines Aufbaues und Entwicklungsgeschwindigkeit
erfordern würde, damit eine rudimentäre Funktionsweise gewährleistet wäre. Dabei
können keine Vergleiche mit 'Riesenformen" von Tieren gezogen werden, die sich manchmal
im Laufe der Evolution entwickelt hatten (Riesenfaultier, Riesenlaufvögel etc.), da
sich in diesen Fällen die Tiere als Gesamtorganismus und über lange Zeiträume verändert
hatten.
Bilder von verschiedenen seltenen Hybriden, die auf natürliche Weise entstanden sind
(inklusive künstliche Besamung), findet man hier:
(http://www.uni-leipzig.de/~mielke/bastarde/bastard5.htm).
Weitere Informationen zu den im Labor erzeugten Schaf-Ziegen Chimären mit Bildern
und Angaben, weshalb es getan wird, finden sie hier:
http://animalscience.ucdavis.edu/faculty/anderson/research.htm.

(c) Shirárch 27.09.2010