| Einführung | Raubtieranatomie | Stammbaum | Molekunargenetik |
| Mutation & Varietät | Genetische Verarmung | Referenzen |
6. Die genetische Verarmung des Gepards und die Konsequenzen
6.1. Der genetische Zustand von Raubkatzen-Populationen
Mit Beginn der in Kapitel 4. angesprochenen, molekularen Ära der Biologie wurden
in den frühen 80er Jahren auch erstmals wildbiologische Fragestellungen mit den
neuen Methoden der Molekulargenetik angegangen. Einige dieser Studien untersuchten
die Frage, welche Faktoren für den Niedergang von bedrohten Tierarten verantwortlich
sind. Stets waren Đberjagung, absichtliche Vernichtung von Beständen, das Ausbleiben
der Beutetiere und die Zerstörung des natürlichen Lebensraumes als Hauptursachen für
das Verschwinden von Tierarten verantwortlich gemacht worden. Zusätzlich wurde jedoch
auch das Zusammenbrechen von Populationen beobachtet, die lediglich geographisch
isoliert, aber zahlenmässig nicht reduziert worden waren (Selander et al., 1973).
Die Arbeitsgruppe um Stephen O'Brien (Laboratory of Genomic Diversity, National
Cancer Inst.,Maryland, USA) publizierte in den 80er Jahren mehrere Studien zur
genetischen Vielfalt bei verschiedenen Raubkatzen (O'Brien et al. 1983, 1985,
1987, 1988).und zog den Schluss, dass beim Gepard als Art eine genetische Verarmung
durch Inzucht vorläge.
Zu den Resultaten im Einzelnen. In der ersten Studie dieser Art wurde die genetische
Variabilität anhand von DNA-Fingerprints (siehe Kapitel 4) ermittelt und es zeigte
sich, dass die Variabilität von Geparden sowohl innerhalb einer bestimmten Population
als auch beim Vergleich verschiedener Unterarten (In Ost- bzw. Südafrika) besonders
gering war. Ein weiterer Test bestand in Hauttransplantationen (die Tiere wurden dazu
zeitweise in Gefangenschaft gehalten).
Schliesslich wurde ein Zusammenhang zwischen der genetischen Verarmung des Gepards
und den folgenden Erscheinungen und Ereignissen hergestellt:
- weltweit abnehmende Populationsgrösse in fragmentierten Verbreitungsarealen
- geringe Spermiendichte bei männlichen Geparden
- ein hoher Prozentsatz abnormaler Spermien
- Ein Fall einer Gepardenkolonie (in Oregon, USA), die durch einen Virus vollständig ausgelöscht wurde.
- hohe Jungensterblichkeit in Gefangenschaft
Die im Fachjournal "Science" vorgestellten Resultate (O'Brien, 1985) stiessen
unter Populationsgenetikern und Ökologen auf einigen Widerstand. Die Hauptaussage
O'Briens, dass die genetische Variabilität der heutigen Geparden für ihr drohendes
Verschwinden mitverantwortlich sei, hat für Artenschutzprogramme wichtige
Konsequenzen. Zudem konnte befürchtet werden, dass in Zukunft weniger Mittel in
Programme für genetisch verarmte Tierarten eingesetzt würde, weil diese Arten
vorzeitig als verloren betrachtet werden könnten.
Andere Studien machten geltend , dass die Fruchtbarkeit des Gepards weniger
eingeschränkt sei, als von den genetischen Analysen erwartet werden könnte, und
dass die schlechten Zuchterfolge in Gefangenschaft hauptsächlich mit dem mangelnden
Verständnis der Ökologie und des Gepardenverhaltens (spezielle Eigenschaften der
Partnerwerbung, soziale Aspekte) zu erklären seien (Caro und Laurenson, 1995).
Zudem wurden die Methoden der O'Brien-Gruppe kritisiert, da in der 1985er Studie
keine anderen Tiere als eine ausgewählte Gepardenpopulation auf Abstossung der
Hauttransplantate getestet wurde. Dieser Punkt wurde 1996 mit einer sorgfältigen
Untersuchung von Hauttransplantat-Abstossung und genetischer Variabilität an
"Pocket-Gophers" (Taschenratten, Thomomys bottae) weitgehend geklärt (Sanjayan
und Crook, 1996). Dabei zeigte sich ein signifikanter Zusammenhang zwischen
genetischer Variabilität, wie sie durch DNA-Fingerprints ermittelt wurde, und
den Resultaten der Transplantat-Abstossung. Somit wurde die Methode der O'Brien-Gruppe
nachträglich validiert.
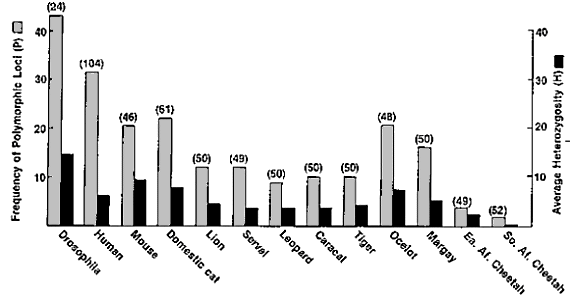
Auf dieser Illustration (5, O'Brien et al., 1987) sind zwei
Masszahlen (Frequenz von polymorphischen Gen-Loci und durchschnittliche Heterozygosität)
für die genetische Vielfalt verschiedener Lebewesen und vor allem Katzen dargestellt.
Rechts aussen sind die Resultate für den Ostafrikanischen (Ea. Af. Cheetah) und
den Südafrikanischen (So. Af. Cheetah) Gepard aufgetragen. Beachten sie auch die
Werte der Hauskatze (Domestic cat), die wesentlich höhere Werte als der Gepard
aufweist.
6.2. Gründer-Effekt: Entstehungsweise und Beispiele
Der Gründer-Effekt (engl. "founder effect") bezeichnet dasselbe Phänomen wie der
Begriff "Flaschenhalseffekt", wenn gleich die Entstehungsweise mit dem ersten
Terminus besser und allgemeingültig illustriert wird: Eine Population von
Individuen (Tiere oder Pflanzen), die zur selben Art gehören, entsteht aus wenigen
Einzelindividuen, die als Gründer der Population wirken. Es gibt grundsätzlich zwei
Entstehungsweisen.
Entweder lässt eine rasche Dezimierung einer ursprünglichen Population (z.B. durch
Krankheiten oder Naturkatastrophen) nur noch wenige Individuen zurück, oder eine
geringe Anzahl Tiere wird geographisch von einer grösseren Ursprungspopulation
isoliert und gründet eine neue Population. Beide Entstehungweisen haben sich in
der Erdgeschichte immer wieder ereignet. Während der erste Mechanismus in der
vorgeschichtlichen Vergangenheit schwieriger nachzuweisen ist, findet man oft
Beispiele für eine geographische Isolation.
Dies findet oft dann statt, wenn eine Insel erstmals von einer bestimmten Tierart
besiedelt wird (Beispiel Galapagos-Vulkaninseln, die einst steril entstanden sind).
In diesem Jahrhundert wurde mehrmals durch menschlichen Eingriff eine Tierart in
ein Gebiet eingebracht, in dem diese zuvor nicht existierte (Beispiel Gir-Forest
in Indien, Ngorogoro-Krater in Afrika). In einigen Fällen ist diese Besiedlung
auf ein einziges Paar der betreffenden Tierart zurückzuführen und die Freisetzung
ist historisch exakt dokumentiert. Zu beachten ist, dass bei diesen Beispielen
keine Zuchtwahl durch Menschen (selten Abschüsse) oder regelmässige Fütterung
etc. stattfand.In solchen Fällen ist es heute möglich, die genetische Variabilität
der Tiere festzustellen, die Effekte des Gründereffekts quantitativ nachzuweisen
und mit grösseren und älteren Populationen derselben Tierart zu vergleichen.
Diese Resultate führten zur Annahme zweier Populations-Engpässe mit nachfolgender,
erneuter Expansion der überlebenden Population des Gepards (Acinonyx jubatus).
Der eine Engpass müsste dabei vor der Aufspaltung der Afrikanischen Unterarten
in den Ostafrikanischen Gepard (Acinonyx jubatus raineyi) und den Südafrikanischen
Gepard (Acinonyx jubatus jubatus) stattgefunden haben (spätes Pleistozän, vor ca.
10'000 Jahren), der andere Engpass nach weiterer geographischer Isolation inklusive
menschlicher Einflussnahme (Dezimierung, Vertreibung, Bewirtschaftung der offenen
Grasländer) in den letzten zweihundert Jahren (Menotti-Raymond et al, 1993).
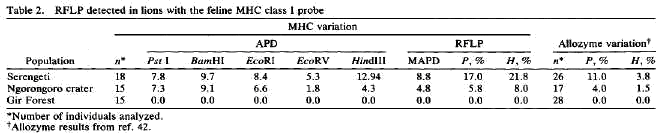
In obiger Tabelle (6, aus Yuhki and O'Brien, 1990) wurden DNA-Fingerprints in
verschiedenen Populationen von Löwen (Panthera leo) ermittelt. Beachten sie bitte
die Schlussresultate für H (Heterozygosity), RFLP (restriction fragment lenght
polymorphism) und Allozymvariation rechts in der Tabelle. Auffällig ist der
eklatante Unterschied zwischen der Variabilität einer Population von Löwen in
der Serengeti, verglichen mit asiatischen Löwen (Panthera leo persica) aus dem
"Gir Forest", einem Schutzgebiet in Indien, in dem nur wenige Paare von Löwen
(ca. 15 asiatische Löwen um das Jahr 1900) nach Đberjagung übriggeblieben waren.
Der Gründer-Effekt bei Raubtieren konnte hier also in einem "Experiment in freier
Wildbahn" beobachtet werden (Chellam et al., 1993).
Weitere Beispiele für den Gründereffekt und resultierende genetische Verarmung:
Als Resultat genetischer Analysen (Roelke et al., 1994) wurde gefunden, dass die noch in der Natur überlebenden 'Florida-Panther' (eine Unterart des Pumas) einen substantiellen Verlust an genetischer Variabilität durch Inzucht zu beklagen haben. Dabei wurde eine geringere genetische Variation als in jeder anderen Puma-Unterart gefunden. DNA-Fingerprints zeigen für diese Pumas eine noch geringere Variabilität als bei den afrikanischen Geparden und beinahe so geringe Werte, wie bei den Löwen des Gir-Nationalparks in Indien. Die Gründe dafür liegen in der sehr weit fortgeschrittenen Aufsplitterung der Lebensräume und daraus folgender Inzucht der Tiere in den übriggebliebenen Habitaten (Culver et al, 2000).
Eine laufende Studie untersucht eine Population von nur noch ca. einhundert Tieren der Iriomote-Wildkatzen (Felis iriomotensis) auf der Insel des gleichen Namens (Präfektur Okinawa, Japan). Die Analyse von Mikrosatelliten-DNA (Kapitel 4.4) zeigte eine so geringe genetische Variabilität, dass mit inzuchtbedingten schädlichen Auswirkungen auf die verbleibende Population in diesem isolierten Insel-Habitat gerechnet werden muss.
(http://www.nies.go.jp/kenko/biotech/bioehp/wildlife.html)
Wie diese Beispiele zeigen, kommt es immer dann zu einer genetischen Verarmung einer Population, wenn die Anzahl Tiere abnimmt und die Grösse des Habitats schrumpft, bzw. fragmentiert wird. Bedingt durch die genetische Verarmung kann es innert kurzer Zeit zu einem Zusammenbruch der Population kommen. Diese Mechanismen sind gut untersucht und auch seit vielen Jahrzehnten bekannt. Es bleibt zu hoffen, dass die Resultate der Genetik weiterhin dazu benutzt werden, die genetische Vielfalt von den in der Natur lebenden Populationen zu verbessern (evtl. mittels Auswilderung aus Tierparks), und nicht nur für nutzlose Zuchtexperimente (Klonen von Geparden, Hybride)!

(c) Shirárch 27.09.2010